









35亿年前,伴随蓝藻的诞生,光合作用登上了历史舞台,并牢牢占据着生物化学循环的C位。植物为了能最大程度的进行光合作用,可谓是耍尽花招,争奇斗艳。究其根本其竞争的本质是对光合作用的三要素:光、水和CO2的竞争。
从广袤的海滨到无边的荒漠,从炎热的赤道到冰封的两极,陆生植物均在使用大气中的CO2。近几个世纪的人类活动使大气层中的CO2浓度从工业革命之前的280 ppm增加到了现在的379 ppm(IPCC2007),但是大气中CO2的变化的程度与水体中一昼夜的变化相比实则小巫见大巫。
自然水体中无机碳的存在形式主要有CO2, HCO3-, CO32-,处于主导地位的无机碳形式由pH调控(图1)。

图1 水体不同 pH 下无机碳存在的形态(Osmond, Winter et al. 1982)
池塘和湖泊中,早晚的pH变化范围为7.0-9.0, 那么CO2在一天之中的浓度可以相差百倍。此外,CO2在水体中的扩散速率仅仅是空气中的万分之一,这进一步限制了沉水植物光合作用时CO2的有效供应。为了适应水体无机碳环境,沉水植物都做了哪些努力呢?
削足适履——水生植物叶片结构的变化
地球上的生命皆诞生于海洋中,植物为了“登陆”,早在4亿年前就演化出了气孔,可以控制大气中CO2的摄入及自身水分的散失。然而现存的沉水植物大多数都没有(功能性的)气孔,导致它们无法通过控制气孔的开合来调节CO2的吸收。沉水植物的叶片通常会发育成非常薄的条形、卵圆形、丝状或羽毛状(图2)。

图2 沉水植物的叶片形态:(A)苦草,(B)龙舌草,(C)穗状狐尾藻(付文龙拍摄)
据统计,全球沉水植物的叶片平均厚度约为130 μm,也就是大约两到三个细胞的厚度,而陆生植物叶片的平均厚度约240 μm(Maberly & Gontero,2018)。这样的叶片结构能增加比表面积,使叶片细胞能最大程度的和水体接触,获取水体中的无机碳。
广开财路——利用底泥中的CO2
水生生态系统中的植物凋落物和动物的尸体残渣有很大一部分会沉积到底泥里,这些富含碳的有机物会在底泥微生物的作用下被分解,产生CO2和CH4。有些沉水植物,在根部和叶片中有大量连续的空洞,它们为CO2从沉积物扩散到叶片提供了路径,从而可以从根部吸收底泥中的CO2,供给植物的光合作用。比如半边莲属的Lobelia dortmanna和水韭属的Isoetes australis 就可以从根部运输底泥CO2到叶片进行光合作用(图3)。

图3 Lobelia dortmanna和Isoetes australis照片(图片来源于网络)
这类植物通常都根系发达、植株矮小,因为CO2从根部大叶片的扩散速率限制了植物的大小(Pedersen et al. 2011)。目前已知的沉水植物种,这类植物占3%左右。
变废为宝——利用HCO3-
自然水体pH通常在7到9之间,在这个范围内,水体中无机碳主要以HCO3-的形式存在(图1)。对于那些只能利用CO2的沉水植物而言,这部分无机碳就是无法利用的废物。但看似“废物”的HCO3-,却被一部分植物当作香饽饽。目前已知的沉水植物中就有约50%的物种可以利用HCO3-作为碳源进行光合作用。
这些植物利用HCO3-的方式也多种多样,有的植物向细胞外分泌碳酸酐酶,将HCO3-催化生成CO2,然后再吸收进叶片;有的植物向胞外分泌H+, 这些H+与HCO3-结合生成CO2,再被吸收;有些植物在细胞膜上演化出了碳酸氢根和阴离子交换通道,直接吸收HCO3-。当然这几种HCO3-利用方式并不是互斥的,它们可以在同一个植物中同时存在(图4)。
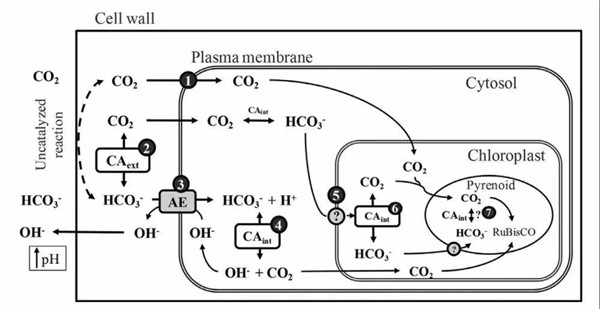
图4 巨藻(Macrocystis Pyrifera)及其利用HCO3-的示意图(Fernandez, P. A. et al 2014)(图片来源于塔斯马尼亚大学)
*Tips1:由于沉水植物吸收HCO3-需要额外的消耗能量,因此沉水植物中HCO3-利用现象虽然普遍,但是并不是在所有条件下都是最优的方式。
节省开支——C4代谢
夏日的正午阳光强烈,光合作用旺盛,水体中的CO2被消耗殆尽,产生大量的O2,导致水体的溶解氧处于饱和状态。在低CO2和高O2的情况下,原本主要起固定CO2作用的酶Rubisco,反而会吸收O2产生CO2,这个过程叫做光呼吸。在光呼吸循环中,有大量的(75%)无机碳会被浪费。为了弥补Rubisco的缺陷,减少无机碳的浪费,沉水植物C4代谢途径应运而生。在C4代谢过程中,从外界吸收的低浓度的CO2由催化效率更高的PEPC固定为含有4个碳原子的酸(比如:草酰乙酸,苹果酸,天冬氨酸等),然后这些酸进入叶绿体,在Rubisco附近释放出高浓度的CO2,抑制光呼吸过程。C4代谢就起到了减少无机碳浪费,并提高光合作用效率的作用。
目前主要在水鳖科植物中发现具有C4代谢过程的沉水植物,包括黑藻(Hydrilla verticillata)、水蕴草(艾格草Egeria densa)、龙舌草(Ottelia alismoides)和海菜花(Ottelia accuminata)等(图5)。


图5 黑藻和海菜花照片(付文龙拍摄)
*Tips2:PEPC以三个碳原子的酸为底物,固定一个CO2,形成四个碳原子的酸,在这个循环中几乎所有的代谢中间物都含有4个碳原子因此称为C4代谢途径。
错位竞争——CAM代谢
既然白天各种水生植物都在争夺水体中的CO2或者HCO3-,且晚上还有呼吸作用产生的CO2,夜晚水体中含有丰富的CO2,那植物为什么不晚上吸收CO2储存起来呢?聪明的沉水植物当然也想到了这点,因此它们也演化出了和陆生植物类似的景天科酸代谢(CAM)途径。CAM途径是在夜间水体中CO2浓度较高的时候吸收二氧化碳,转变为有机酸(一般为苹果酸、柠檬酸或者天冬氨酸等)并存储在液泡中。白天这些酸被运输到叶绿体,释放出CO2供给光合作用。这个过程也就导致了植物细胞在夜间的酸度比白天要高。
最典型的CAM代谢水生植物就是中华水韭(Isoetes sinensis),其昼夜酸度差可以高达110μequ/g鲜重(Yin et al2017 )。除了中华水韭之外,泽番椒(Deinostema violaceum)也具有较强的CAM代谢能力,刺苦草也可能具有CAM途径。

水韭(图片来源:中国植物志)
*Tips3:CAM,景天科代谢,最初在景天科植物中发现他们细胞内的酸度在昼夜有很大的变化,因此命名为“景天科-酸-代谢”(Crassulaceanacid metabolism)。现在许多教材或文献中都被误解为“景天酸代谢”,事实上并没有一种叫“景天酸”的物质。
多管齐下——多途径碳固定
所谓武林高手,当然不止会一门功夫。为了更好的吸收无机碳进行光合作用,沉水植物当然也不止有一种吸收和固定无机碳的途径。比如龙舌草叶片只有两三层细胞,但它既可以吸收CO2,也可以利用HCO3-,还具备C4代谢途径,在低CO2环境下还能发展出CAM代谢,可谓集多种碳浓缩机制于一身。
正是因为沉水植物的种种努力,才换来了它们在全球碳循环中不可或缺的地位。据估计海洋沉水植物固定的碳为40Pg每年,大约贡献了全球海洋碳埋藏的20%,对维持全球碳库的周转和稳定起到了重要作用。
参考文献:
1. Fernandez PA,Hurd CL, Roleda MY.(2014).Bicarbonate uptake via an anion exchange protein is the main mechanism ofinorganic carbon acquisition by the giant kelp Macrocystis pyrifera (Laminariales,Phaeophyceae) under variable pH. J Phycol 50(6): 998-1008.
2. Maberly,Stephen Christopher; Gontero, Brigitte. (2018). Trade-offs and synergies in thestructural and functional characteristics of leaves photosynthesizing inaquatic environments. In: The leaf: a platform for performing photosynthesis.Springer, Cham, 2018. p. 307-343.
3. Osmond, C.(1978). "Crassulacean acid metabolism: a curiosity in context."Annual Review of Plant Physiology 29: 379-414
4. Pedersen O,Pulido C, Rich SM, Colmer TD. (2011). In situ O2 dynamics in submerged Isoetesaustralis: varied leaf gas permeability influences underwater photosynthesisand internal O2. J Exp Bot 62:4691–4700
5. Solomon S.2007. IPCC (2007): Climate change the physical science basis. AGUFM 2007:U43D-01.
6. Yin L, Li W,Madsen TV, Maberly SC, Bowes G. (2017). Photosynthetic inorganic carbonacquisition in 30 freshwater macrophytes. Aquatic Botany 140: 48-54
撰文:江红生



